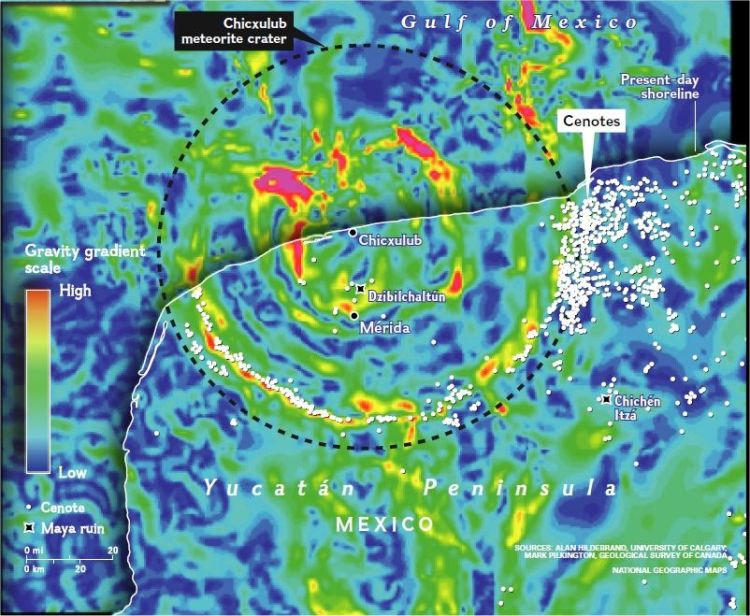
“Lucifer’s Hammer killed the dinosaurs,” said US physicist Luis Alvarez, in a lecture on the geochemical evidence he and his son found of a massive impact at the end of the Cretaceous period. A year later, Pemex (a Mexican oil company) identified Chicxulub as the site of this massive asteroid impact. The crater is more than 180 km (110 miles) in diameter and 20 km (10 miles) in depth. The impact released an estimated energy equivalent of 100 teratonnes of TNT, induced earthquakes, shelf collapse around the Yucatan platform, and widespread tsunamis that swept the coastal zones of the surrounding oceans. Global forest fires might have raged for months. Photosynthesis stopped and the food chain collapsed. The decrease of sunlight caused a drastic short-term global reduction in temperature. This phenomenon is called “impact winter”. Cold and darkness lasted for a period of years. Three-quarters of the plant and animal species on Earth disappeared, including non-avian dinosaurs, pterosaurs, marine reptiles, ammonites, and planktonic foraminifera.
Early work speculated that the eruption of the Deccan Traps large igneous province was the main abiotic driver of the K/Pg mass extinction. However, in the late ’70, the discovery of anomalously high abundance of iridium and other platinum group elements in the Cretaceous/Palaeogene (K-Pg) boundary led to the hypothesis that an asteroid collided with the Earth and caused one of the most devastating events in the history of life.
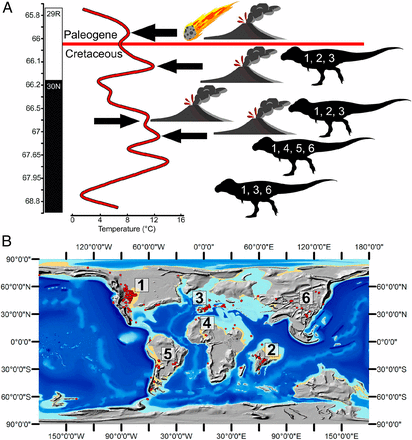
Geologic (A) and paleontological (B) records of the K/Pg mass extinction. From Chiarenza et al., 2020.
The Deccan Traps in central India is formed from a series of short (∼100-ky) intermittent eruption pulses, with two main phases: one toward the end of the Cretaceous, and the other starting just before the boundary and continuing through the earliest Paleogene. A new study from Imperial College London, the University of Bristol and University College London, lead by Dr Alessandro Chiarenza, compared the climatic perturbations generated by Deccan volcanism and the asteroid impact. The new study found that the extreme cooling caused by the asteroid impact created the conditions for the dinosaur extinction worldwide. Additionally, they found that the Deccan’s influence after the event might have been of greater importance in determining ecological recovery rates after the asteroid-induced cooling, rather than delaying it.
Previous studies suggested that while the surface and lower atmosphere cooled (15 °C on a global average, 11 °C over the ocean, and 28 °C over land), the tropopause became much warmer, eliminate the tropical cold trap, and allow water vapor mixing ratios to increase to well over 1,000 ppmv in the stratosphere. Those events accelerated the destruction of the ozone layer. During this period, UV light was able to reach the surface at highly elevated and harmful levels.
References:
Alfio Alessandro Chiarenza, Alexander Farnsworth, Philip D. Mannion, Daniel J. Lunt, Paul J. Valdes, Joanna V. Morgan, and Peter A. Allison. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. PNAS, 2020 DOI: 10.1073/pnas.2006087117
P.M. Hull et al., “On impact and volcanism across the Cretaceous-Paleogene boundary,” Science (2019). Vol. 367, Issue 6475, pp. 266-272 https://science.sciencemag.org/content/367/6475/266
Alvarez, L., W. Alvarez, F. Asaro, and H.V. Michel. 1980. Extraterrestrial cause for the Cretaceous-Tertiary extinction: Experimental results and theoretical interpretation. Science 208:1095–1108.









